Reading time: 6 minutes
Limitation #1 - Physical properties
The only physical property that EEG detects is voltage.
Because voltage is defined as the difference in electric potential, 2 points of measurement are required to measure voltage. The implications of this are as follows:
- Every EEG electrode has to be compared to one other electrode in order to measure voltage. There’s 2 methods of selecting those “reference” electrodes:
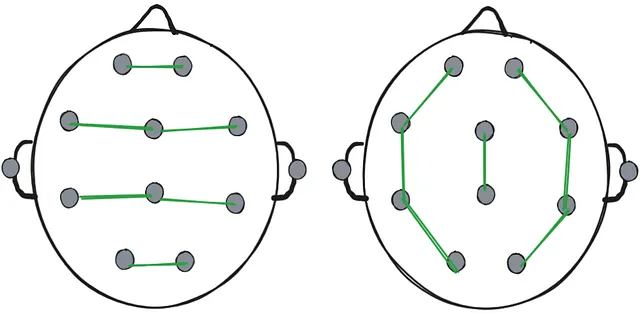
- “Referential montage”: One particular electrode serves as a reference to all others. The chosen electrode is usually the one least affected by the activity from the brain, such as an electrode at the ear.
- “Bipolar montage”: All electrodes are paired up with another electrode next to them forming “chains”.
In most cases both montages are recorded and the person analysing the EEG recording can switch between them.
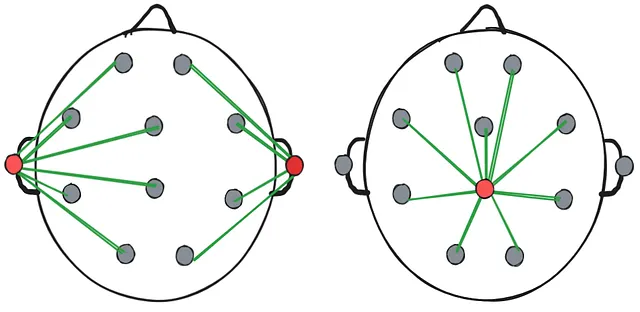 Two examples of a referential montage. Left: the reference electrodes are at the ears. Right: the reference electrode is the central electrode.
Two examples of a referential montage. Left: the reference electrodes are at the ears. Right: the reference electrode is the central electrode.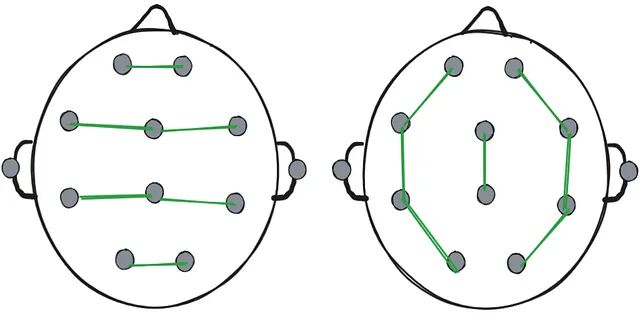 Two examples of bipolar montage. Left: the electrodes are chained horizontally. Right: the electrodes are chained vertically.
Two examples of bipolar montage. Left: the electrodes are chained horizontally. Right: the electrodes are chained vertically. - It’s not possible to measure “absolute voltage” or “absolute strength of the brain signal” at any one electrode - each measurement is a relative difference in electric potential between 2 points. This means that if there’s high-strength activity spread out through a large area of the brain that includes the reference electrode, voltage picked up by EEG in that area would be low as the relative difference between electrodes is low.
Some technologies that can measure different physical properties of the brain are as follows:
- MEG: detects magnetic field. As changing magnetic field creates an electric field at 90 degree angle to it and vice versa, MEG and and EEG are often used together to detect activity that the other is less sensitive to due to angles at which the activity propagates.
- fMRI: detects changes in blood oxygenation level.
- PET: detects movement of radioactive traces injected into the bloodstream.
- fNIRS: detects blood flow and oxygenenation level using infrared light.
- MRI: measures tissue density based on how long hydrogen nuclei continue to emit energy.
- CAT: measures tissue density based on how much of X-ray waves is absorbed.
Limitation #2 - Number of neurons
The skull is a poor conductor of electricity. A single neuron generates around 70-100 mV and over 99% of that is lost passing through the skull. While the EEG can detect voltage on the scale of μV, the amount that reaches the top of the skull is indistinguishable from background noise (electromagnetic fields generated by muscle activity, electronic devices etc.). To distinguish itself from background noise, a large number of neurons need to fire simultaneously - at least 10,000 - 50,000.
This is not a large number when it comes to neurons, as one cubic milimetre of the cortex contains around 50,000 - 150,000 neurons. While a small number of neurons (1-10) is often enough to trigger or detect something (e.g. trigger a memory or detect a specific odor molecule), most activity involves networks of millions of neurons - even simple activity such as moving a finger.
Limitation #3 - Area and depth
Because the signal from the neurons is “blurred” while passing through the skull, it’s not possible to pinpoint the exact area the signal is coming from - the spatial resolution of EEG is around 6-10 cm^2.
As EEG is located on top of the scalp and brain activity is less detectable the further away it is, EEG is dominated by cortical surface activity and doesn’t usually pick up the activity from deep brain regions such as the hippocampus or the thalamus. This activity can be detected with the use of some machine-learning algorithms (e.g. HEAnet) or high-density EEG (64+ electrodes).
Furthermore, the EEG is generally not good at locating the depth of the activity - a deep source can produce the same pattern as a broad shallow source. Therefore, EEG can’t easily identify the specific region of the brain the activity is coming from. This is a well-known unsolved problem in neuroscience - “the inverse problem”:
-
The inverse problem: Given the voltages measured on the scalp, what neural sources inside the brain produced them? This problem is impossible to solve, because infinitely many internal configurations can produce the same scalp pattern.
-
The forward problem: Given the neural sources inside the brain, what voltages are measured on the scalp? This problem can be solved, as once all the variables are known (geometry of the head, conductiveness of tissues, electrode positions etc.), the physics is deterministic - these variables can be plugged into the equation for electric potential and the equation produces one solution.
Several algorithms exist to localise the source of EEG activity given the voltage measured on the electrodes, and all of them depend on the forward problem - knowing how various activity translates to voltage measured on the scalp, they can predict the most likely source given the measured activity.
Limitation #4 - Direction
Over 70% of neurons in the cortex (the outer layer of the brain) are pyramidal neurons. Pyramidal neurons are oriented perpendicularly to the brain surface, as such:
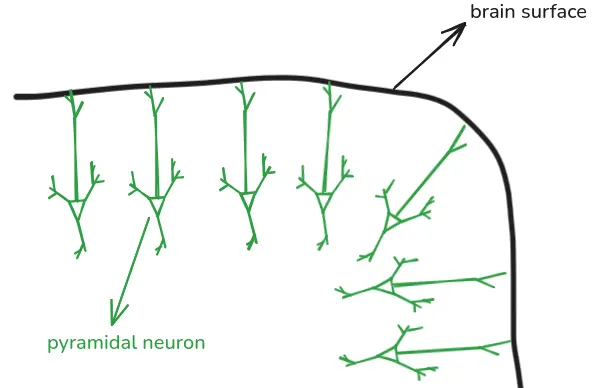
The surface of the brain is folded - parts of the surface are either “hills” (called gyri) or “valleys” (called sulci). Due to pyramidal neurons being oriented perpendicularly to the brain surface, the neurons are generally aligned either vertically or horizontally to the scalp. Neurons in gyri are aligned vertically, and neurons in sulci horizontally:
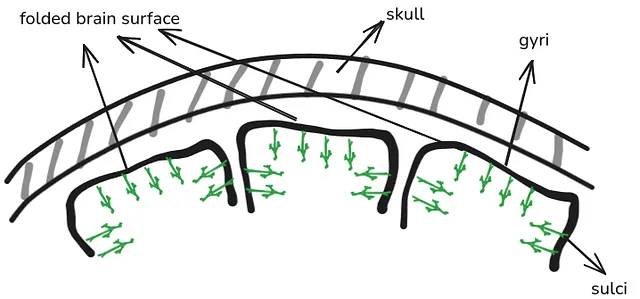
When a neuron is involved in some brain activity, the electric potential difference is oriented along the length of the neuron - either the top of the neuron is positive and the bottom of it negative, or vice versa. Therefore, for neurons aligned vertically to the scalp, the part of the neuron closest to the scalp has a clearly defined positive or negative electric potential. This makes it easy for the potential to be picked up by EEG.
In contrast, for neurons aligned horizontally to the skull, the electric potential difference is oriented sideways. This makes it more “spread out” and more likely to be muddled by the sideways-oriented potential by other neurons around it. This makes it less clearly defined, and much more difficult to be picked up by EEG.
Therefore, the activity that EEG picks up the most is the activity from pyramidal neurons that are oriented perpendicularly to the scalp - that is, neurons in the gyri.
The opposite is true for MEG, which is more sensitive to activity from parallelly-oriented neurons due to the way that the magnetic fields extend outside the head. This is another reason why EEG and MEG compliment each other well.
Limitation #5 - Type of neural activity
Three types of electrical activity are seen for neurons:
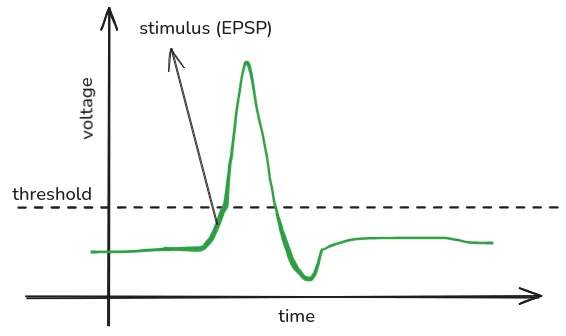
- Action potential (spikes):
- Rapid reversal of electric charge inside the neuron.
- Represents the neuron firing after achieving enough stimulation (once it becomes positively charged enough).
- Lasts around 1 milisecond.
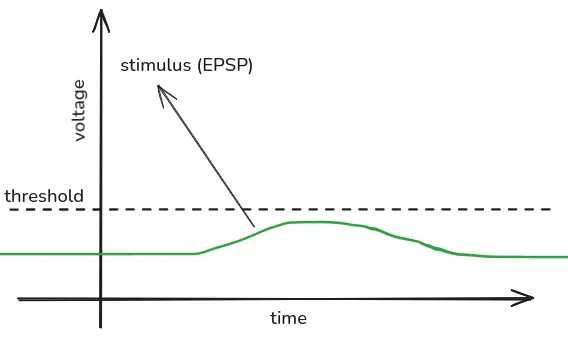
- Excitatory Postsynaptic Potential (EPSP):
- Positive ions flow into the neuron, making the neuron more positive and more likely to fire (“exciting” the neuron).
- Lasts 10-100 miliseconds.
- Inhibitory Postsynaptic Potential (IPSP):
- Negative ions flow into the neuron or positive ions flow out of it, making the neuron more negative and less likely to fire.
- Lasts 10-100 miliseconds.
EEG picks up on EPSP and IPSP, but doesn’t pick up on spikes for the following reasons:
- EPSP and IPSP create a stable electric potential difference (one end of the neuron is more positively/negatively charged that the other end), which can be more easily picked up by EEG. Spikes don’t create a stable potential difference, as they travel along the neuron like a wave.
- EPSP and IPSP last significantly longer than spikes. This makes them more likely to overlap in time with other neuron’s EPSP and IPSP, which will add up and create a large enough field that EEG can detect.